Адаптация
- Информация о материале
- Категория: Объёмное телевидение – символ XXI века
- Опубликовано: 09.01.2015 14:51
- Автор: Павел Варгин
Мы живем в мире, наполненном светом. Диапазон изменения яркостей окружающих объектов может достигать пятнадцати порядков (квадриллиона раз). Это диапазон изменения значений пленоптической функции, о которой мы говорили в начале статьи. На рис. 17 приведен диапазон яркостей белой бумаги освещаемой различными источниками освещения. Даже создание такого диапазона яркостей в экспериментальных условиях представляет известную проблему. Во всем этом диапазоне глаз сохраняет работоспособность, хотя как жаловался Ньютон (1691) в своем письме Локку, ему пришлось трое суток сидеть в полной темноте, чтобы избавиться от послеобразов Солнца после наблюдении его в телескоп. Бойль (1664) в таких опытах получил ожог сетчатки и продолжал видеть образ Солнца с закрытыми глазами даже спустя 10 лет. С другой стороны после нескольких часов проведенных в темноте человек может видеть ничтожную вспышку света эквивалентную восприятию десяти фотонов. Ни одна телевизионная камера не может перекрыть такой диапазон яркостей. В лучшем случае мы можем попытаться перекрыть его с помощью трех телекамер разных диапазонов, охваченных глубокими параметрическими обратными связями автоматических регулировок режимов (коэффициентов усиления).
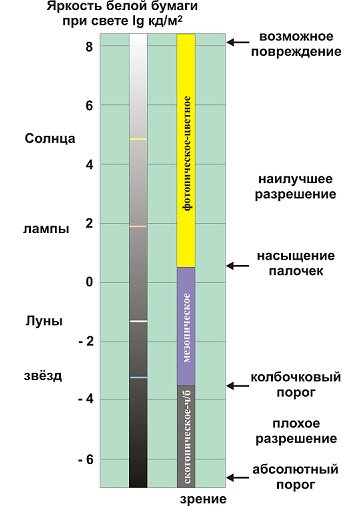
Рис. 17. Диапазон яркостей белой бумаги при различных источниках освещения.
При использовании трехмерной интерактивности объёмного телевидения можно представить себе следующие ситуации: сначала вы находились в помещении, а затем вышли из дома полюбоваться закатом или, двигаясь в автомобиле по ярко освещенной автостраде, заехали в тоннель. Перепады яркости, вызванные вашим поведением, весьма велики и для реалистичности объёмного изображения телевизионная модель должна содержать информацию о текущей освещенности на местности.
В последние годы вызывает большой интерес проблема отображения тона (TM - ToneMapping ) – сжатия светового диапазона содержащегося в изображении широкого динамического диапазона (HDR – HighDynamicRange) в визуализируемый узкий динамический диапазон (LDR – LowDynamicRange). Например, в Минесотском Институте прикладной математики (2009) разработан алгоритм ТМ на основе модифицированного уравнения Нака – Раштона, позволяющий не только сжимать HDR, но и осуществлять локальное повышение контраста изображения и устранять потери цвета.
Необходимо отметить, что разброс коэффициентов отражения поверхностей природных объектов не велик, обычно не более 40 раз. Примерно такой же диапазон яркости отображает зрение человека без адаптации. Классический пример отображения связан с чтением книги. В помещении текст нам кажется черным, а лист книги белым. На ярком солнечном свете текст, все равно нам кажется черным, а лист белым, хотя объективно яркость текста на солнце в десять раз больше яркости листа в помещении. Учитывая такие адаптационные свойства зрения достаточно иметь монитор с тысячекратным изменением яркости для воспроизведения любых изображений, в том числе и с имитацией пересветки. Современные цифровые фотокамеры обладают большим динамическим диапазоном и фиксируют HDR в RAW(англ. raw — cырой) файлах (до 14 бит на канал) или Tiff(англ. Tagged Image File Format) файлах (до 64 бит на канал) . Для сравнения в широко используемом стандарте Truecolor яркость (цвет) кодируется всего 256 уровнями (8 бит на канал), при этом глаз не ощущает «ступенек» тона. Обладая запасом разрешения по градациям яркости можно растягивать при воспроизведении диапазон яркости малоконтрастных локальных участков, повышая их локальный контраст.
Благодаря процессу эволюции, подвергаясь непрерывным колебаниям освещенности при смене дня и ночи, мы получили механизм зрения, обладающий феноменальными динамическими характеристиками. Обычно говорят об адаптационной роли зрачка глаза (изменение площади в 16 раз) и подчеркивают наличие двух видов фоторецепторов сетчатки глаза: палочек и колбочек, которые делят диапазон воспринимаемых яркостей на два. В действительности основная «тяжесть» адаптационной перестройки падает на биохимический комплекс механизма трансдукции фоторецепторов (изменение передачи в миллионы раз), а также на нейросетевой механизм организации рецептивных полей сетчатки глаза.
Механизм фототрансдукции обеспечивается длинным каскадом биохимических реакций, заканчивающийся цГМФ (циклический гуанин моно фосфат) регуляцией пропускания мембранных каналов рецепторов. Изменение ионной проводимости мембраны фоторецептора вызывает изменение ее внутриклеточного потенциала (минус 35 мВ). Увеличение яркости света вызывает гиперполяризацию мембраны, до минус 70 мВ, уменьшение - деполяризацию, до 0 мВ. Градуальный потенциал рецепторов воздействует на биполярные и горизонтальные нейроны сетчатки глаза и далее на ее амакриновые и ганглиозные нейроны так, что у ганглиозной клетки образуется приблизительно круглое рецептивное поле, с выраженными центром и переферией.
Схема восприятия света для дневного (колбочкового) зрения показана на рис. 18. Три вида колбочек сетчатки глаза, изображены разными цветами в соответствии с их спектральными характеристиками: L (длинноволновые) – красным, M (средневолновые) – зеленым и S (коротковолновые) - синим. Каждая из колбочек обладает собственной системой автоматической регулировки усиления (АРУ).
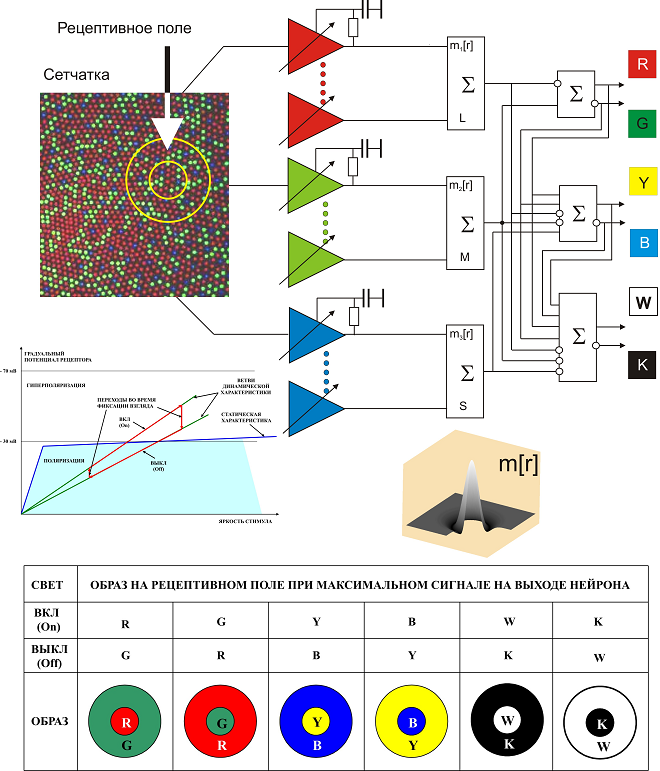
Рис. 18. Функциональная схема восприятия света первичного уровня зрительной системы. Вейвлет «сомбреро» (с вырезанной для наглядности четвертью) представляет собой весовые функции трех сумматоров сигналов колбочек концентрических рецептивных полей (РП) нейронов. Вид характеристик АРУ колбочек РП соответствует раздражению мигающим светом. Снизу приведена таблица реакций нейронов на различные образы.
На схеме это показано с помощью усилителей, изображенных треугольниками, охваченных цепями параметрической обратной связи, воздействующей на коэффициент их усиления. Запаздывающий характер этой связи изображен с помощью интегрирующих RCцепочек. Постоянная времени АРУ колбочек составляет величину порядка нескольких секунд. Роль обратной связи в механизме фототрансдукции, как считают, выполняют в основном ионы кальция, воздействуя на различные ее фазы. Вид статической и динамических характеристик АРУ колбочки, приведенный на рисунке, соответствует периодическим скачкам взгляда с яркого на темный фрагмент объекта наблюдения и обратно. Важной особенностью усилителя с АРУ является различие его коэффициентов усиления для низких и высоких временных частот, что соответствует различным наклонам его статической и динамической характеристик.
Сигналы колбочек каждого вида с площади, очерченной большим кругом рецептивного поля, поступают на соответствующий сумматор. Каждый сумматор осуществляет взвешенное суммирование сигналов колбочек. Весовые функции сумматоров, можно аппроксимировать двумерными вейвлетами типа «сомбреро» (mexhat). Положительная (возбудительная) часть вейвлета соответствует внутреннему (малому) кругу рецептивного поля, а отрицательная (тормозная) его внешнему кольцу. Здесь центр слегка преобладает над периферией в отличие от принятого в вейвлет анализе баланса, что позволяет фиксировать постоянную составляющую сигнала. Выходные сигналы сумматоров обрабатываются согласно приведенной схеме. Небольшие белые кружки на входах и выходах элементов схемы обозначают инверсию сигналов. Результат обработки представляется сигналами на шести выходах: R (red - красный), G (green - зеленый), Y (yellow - желтый), B (blow– синий), W (wait – белый), и K (black - черный. Эти выходы соответствуют шести отдельным нейронам, с одинаковыми по площади, местоположению и форме рецептивными полями. Выходные сигналы здесь положительные (соответствуют деполяризации нейрона) и могут присутствовать либо на выходе R, либо на выходе G, либо отсутствовать на обоих. Аналогичное ограничение распространяется на пары Y, B и W, K. Такую организацию распределения выходных сигналов принято называть оппонентной. Согласно таблице внизу рис. 18, при проецировании на рецептивное поле подходящего образа, при включении или выключении света будет получена максимальная реакция (сигнал) соответствующего нейрона. При освещении рецептивного поля однородным по цвету излучением реагирует нейрон, соответствующий цвету излучения.
Важной особенностью работы данной схемы является небольшой коэффициент усиления для сигналов низких пространственных частот и большой для частот, на которые настроен вейвлет. Необходимо отметить, что разбалансировка вейвлета может меняться при увеличении внешней освещенности в сторону уменьшения относительного веса его положительной части, то есть в зрительной системе присутствует еще одна петля обратной связи АРУ не показанная на схеме.
Приведенная схема имеет иллюстративное значение (показывает наличие различных механизмов зрения) и не может приниматься за вполне адекватную модель обработки сигналов зрительной системой человека. Элементы более точной модели можно найти, например, у Ванхатерена (2007), где учитывается действие колбочек, биполярных и горизонтальных клеток сетчатки глаза. В частности там учтен нелинейный характер передачи сигналов в сетчатке. Наша схема соответствует оппонентной теории цветовосприятия, предложенной Эвальдом Герингом в ее линейном приближении. Учет нелинейности, по-видимому, можно свести к утверждению о том, что оппонентные выходы представляют нам не разность, а отношение входных сигналов.
Мы ощущаем красный цвет, когда возбуждаются нейроны R, при более активном действии колбочек L-типа, или ощущаем зеленый цвет, когда возбуждаются нейроны G, при преобладании действия колбочек M-типа, либо не ощущаем этих цветов при балансе действия этих типов колбочек, когда не Rне G не возбуждаются.
Мы ощущаем желтый цвет, когда возбуждаются нейроны Y, при преобладании суммарного действия колбочек L и M типов над действием колбочек S-типа, или ощущаем синий цвет, когда возбуждаются нейроны B, при преобладании действия колбочек S-типа, либо не ощущаем этих цветов при балансе действия этих типов колбочек. Обратите внимание на то, что по схеме суммарное действие колбочек L и M типов берется без значения разности их действия.
Мы ощущаем степень светлоты объекта наблюдения, когда срабатывают нейроны W или степень его затененности, когда срабатывают нейроны K, по сравнению со средним уровнем ощущения яркости сцены.
Градуальные выходные сигналы схемы в реальной сетчатке преобразуются ганглиозными клетками в частотно-импульсные сигналы потенциалов действия и по аксонам, которые составляют зрительный нерв, передаются в головной мозг.
Снижение удельного веса нижних пространственных частот приводит к эффекту хорошо видному на тесте Э. Эдельсона (рис.19), где клетки А и В, а также соединяющая их линия, кажутся различными по яркости, хотя на самом деле таковыми не являются.

Рис. 19. Тест Эдельсона.
Такая организация преобразователя свет-сигнал, которую нам демонстрирует зрительная система человека, заслуживает пристального внимания с технической точки зрения, поскольку речь идет о возможности резкого сокращения динамического диапазона светоприемника.
Отметим, что весовые функции рецептивных полей сетчатки увеличивают свой масштаб по мере удаления от ее центра, что приводит к снижению разрешающей способности сетчатки на периферии (рис. 11). Это соответствует возможности вычисления вейвлет-спектра изображения по осям сдвига и масштабирования, накапливая данные при разнообразных скачках (саккадах) глаз.
С другой стороны во время дрейфа глаз (рис. 13) изображение с небольшой скоростью (до 20 элементов в секунду) смещается по сетчатке глаза. В телевидении используются телевизионные камеры (ИК диапазона), в которых реализовано так называемое оптическое микросканирование (искусственный дрейф) с помощью вращающейся наклонной стеклянной плоскопараллельной пластинки. Благодаря этому техническому приему разрешающая способность телекамер существенно (в несколько раз) повышается. Возможно, что дрейф глаз используется зрительной системой человека на ее высоком уровне с той же целью, преодолевая потерю разрешения на стадии формирования рецептивных полей.
Мы рассмотрели ряд механизмов начальных стадий работы зрительной системы человека. Уже на этих стадиях нас поражают ее возможности парировать гигантские изменения освещенности окружающей среды и добиваться высокого пространственного и временного разрешения при наличии весьма скромных возможностей по быстродействию.
Эволюция биологических и технических систем отображения окружающего мира идет, на первый взгляд, различными путями. Между ними имеется, однако, много общего начиная с подобия генных мутаций и изменчивости технических идей и заканчивая прямым заимствованием молодым, мыслительным эволюционным потоком технических решений у старушки природы. В качестве примера попытки такого заимствования отметим сообщение IBM(2008) о начале реализации проекта разработки принципиально новой архитектуры аппаратной вычислительной системы, строящейся на принципах организации живого мозга. Официальное название проекта – «Создание компьютера, способного к познанию с помощью синаптроники и высокопроизводительных вычислений» («Cognitive Computing via Synaptronics and Supercomputing» (C2S2)). Предполагается, что мозг, скорее, представляет собой не нейронную, а синаптическую сеть, а мышление является результатом биохимической организции мозга.
Сигналы, поступающие в мозг от сетчаток глаз, подвергаются им сложной обработке. На корковом уровне работы зрительной системы, производится выделение направлений контуров и движений объектов наблюдения с помощью сложных рецептивных полей. Имеются данные (И.А.Шевелев, 2007) о том, что, ориентационная чувствительность нейронов зрительной коры динамически перестраивается за время одной саккады с дискретностью в 20 миллисекунд от вертикальной до горизонтальной ориентации. С помощью метода функциональной магнито-резонансной томографии удалось выявить у человека до 36 зон коры мозга, осуществляющих последовательную и параллельную обработку зрительной информации. Считается, что обработка идет по принципу детектирования все более обобщенных признаков распознаваемых объектов наблюдения. Обнаружены механизмы синхронизации процессов обработки в виде волн электрической активности мозга. Изучая мозг, можно видеть, что процесс анализа информации о внешнем мире идет параллельно с ее сжатием.