
Введение
- Информация о материале
- Категория: Оппонентно-цветовое кодирование изображений
- Опубликовано: 19.05.2023 16:44
- Автор: Павел Варгин
При разработке систем технического зрения в рамках проектов по созданию искусственного интеллекта большое значение имеет бионический подход с ориентацией на зрительные системы различных организмов, созданные в процессе их естественной эволюции. Важным свойством зрения является цветоразличение. Если для таких направлений техники, как цветовоспроизведение и колориметрия может быть достаточным представление о цвете как чувстве яркости (количественная характеристика) и цветности (качественная характеристика), то для инженерии искусственного интеллекта и физиологии этого явно недостаточно. Здесь необходимо комплексное представление о зрении, как таковом, и цвете, как его свойстве.
Зрение помогает нам создавать в сознании образ (модель) окружающего мира. Зрение не единственное наше чувство (модальность), а одно из многих, и образ мира в сознании человека многомодален. Каждый из известных органов чувств обладает своими рецепторами (датчиками-преобразователями физических сигналов). Преобразованные сигналы передаются в мозг для формирования динамичного образа мира. Этот образ помогает субъекту
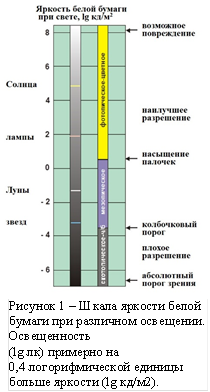
(носителю сознания) реализовать свои цели на основании собственного опыта, планируя будущее. Физическим сигналом воздействия на глаз является свет – небольшой участок электромагнитного спектра волн, характеризуемый яркостью, угловым распределением яркости и формой спектра. Свет в диапазоне длин волн 400…760 нм считается видимым. Если изменение яркости по полю зрения обычно не превышает двух порядков при изменении коэффициентов отражения объектов наблюдения, то яркость видимого излучения может изменяться в гигантском диапазоне, приблизительно от 10–7 до 10+8 кд/м2, как показано на рисунке 1. Здесь можно видеть поддиапазоны работы различных фоторецепторов сетчатки глаза: палочек, работающих при низких освещённостях, и колбочек – при высоких. И те, и другие способны к адаптации в очень широких диапазонах яркости. Адаптация фоторецепторов обеспечивается в основном биохимическим механизмом фототрансдукции [1].
Скорость адаптации высока. Оценить эту скорость можно по скорости исчезновения изображения, предъявляемого глазу, при искусственной стабилизации изображения на сетчатке. Уже через 1 – 2 секунды изображение исчезает даже при стопроцентном контрасте исходного изображения. При этом человек начинает видит сплошное серое, так называемое, «пустое поле» [2].

Препятствуют исчезновению изображения микродвижения глаз, среди которых: микросаккады, дрейф и тремор. Запись микродвижений глаз приведена на рисунке 2. Высоко частотный тремор, (круговое движение ~ 90 Гц) накладывается на медленные дрейфы (изогнутые траектории). Микросаккады – быстрые (~ 25 мс) скачкообразные движения, обычно возвращают образ к центру визуализации (прямые линии). Диаметр части центральной ямки поля зрения, показанной здесь, составляет 0,05 мм (~ 10 угл. мин.).
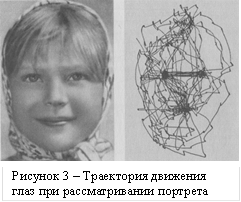
Другие движения глаз – скачки (саккады) необходимой амплитуды используются для направления взора на объекты внимания субъекта. Запись таких движений глаз приведена на рисунке 3. Для слежения за движущимися объектами используются прослеживающие движения глаз (динамическая фиксация) [3]. Запись движения глаз осуществляется с помощью особой техники [2].
Движения глаз имеют определяющее значение не только для перечисленных целей, но и для:
субдискретизации изображения с целью повышения разрешающей способности зрения;
распространения синего цвета на область ясного зрения.
Далее рассмотрим достижение этих двух целей с учётом феномена пространственной константности зрения [4].
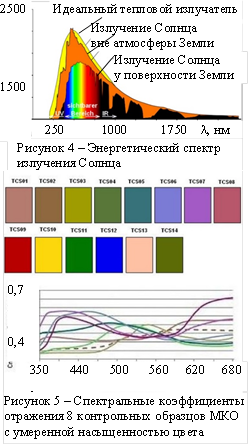
Форма спектра света, отражённого от объекта, зависит как от спектра освещения, так и от спектральных характеристик отражения объекта. На рисунке 4 показан спектр источника освещения – Солнца в единицах Вт/м2 мкм. Заметим, что цветовая температура излучения Солнца обычно не совпадает с температурой идеального теплового излучателя – 5900 К, спектр которого приведён на рисунке, поскольку зависит от фильтрующих свойств атмосферы и положения солнца на небосклоне. На рисунке 5 показаны спектральные коэффициенты отражения контрольных образцов международной комиссии по освещению (МКО). Существуют различные способы оценки формы спектра отражённого излучения. Например, в технике существует направление разработки, так называемых, спектрозональных специальных телевизионных систем, где количество зон или «цветов», по существу, не ограничено. Различать цвета можно по соотношению долей полной световой энергии, прошедших через зональные светофильтры.
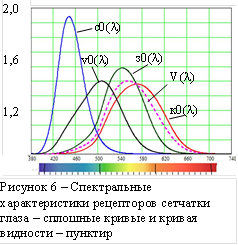
В отличии от зрения собак, кошек или кроликов, зрение большинства приматов и человека обладает цветоразличением. У человека обнаружены четыре типа фоторецепторов, с различными спектральными характеристиками – относительными спектральными чувствительностями. На рисунке 6 приведены эти характеристики (сплошные кривые, вместе с пятой (пунктир), так называемой кривой видности, производной от характеристик колбочек. Характеристики колбочек: коротко- (S, синия), средне- (M, зелёная) и длинноволновой (L, красная), – перекрываются (см. рисунок 6). Оставляя в стороне вопрос о работе палочек и взаимодействии палочек и колбочек, отметим только эффект Пуркинье – смещение кривой видности влево до слияния с характеристикой палочек при снижении освещённости. Далее здесь рассматриваем работу колбочек в условиях фотопического зрения. Абсолютная спектральная чувствительность колбочек не может быть определена из-за упомянутой выше адаптации колбочек в очень большом световом диапазоне. Однако относительная спектральная чувствительность в сравнительно узком диапазоне яркостей может быть определена. Методом уравнивания постоянной энергетической яркости различных по длине волны монохроматических излучений основными излучениями трёхцветного визуального колориметра получают так называемые кривые сложения трихромата (человека с нормальным цветовым зрением), характерные для данного колориметра. Далее, исследуя на этом же колориметре особенности цветовосприятия дихроматов (красно-, зелёно- или даже очень редких сине-слепых), по всем полученным данным определяют кривые относительной спектральной чувствительности [5].
Заметим, что в литературе по колориметрии можно видеть различные кривые сложения с отрицательными значениями цвета, которые соответствуют проекции некоторых из основных цветов колориметра на поле измеряемого цвета, а не на поле слагаемых цветов. Форма кривых сложения зависит также от выбранной цветовой модели: XYZ, RGB и др. Кривые относительной спектральной чувствительности фоторецепторов глаза всегда находятся в области положительных значений чувствительности и не зависят от типа колориметра и модели.
Кривые колбочковых спектральных чувствительностей обычно нормируют по площадям под ними домножением на соответствующие коэффициенты и тогда, при вычислении реакции колбочек на излучение с равномерным энергетическим спектром, эти реакции равны для всех типов колбочек. Одинаковые реакции разнородных колбочек на такой спектр освещения порождают ощущение белого (серого) цвета. Ощущение белого можно также вызвать сочетанием всего трёх первичных цветов колориметра или цветного монитора с характерным для каждого из них отношением яркости этих цветов. Отношения задаются с помощью так называемых яркостных коэффициентов цветов.
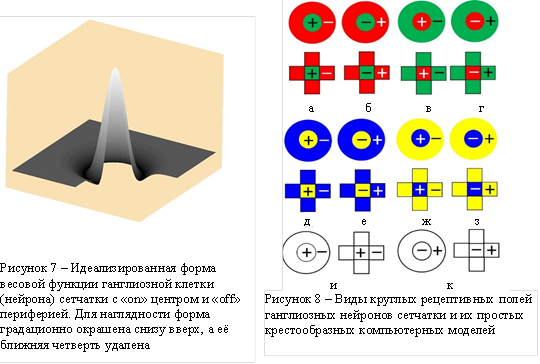
При изучении зрительной системы мы сталкиваемся с кажущимся противоречием. С одной стороны, мы видим полноцветную раскраску окружающих нас объектов, а с другой стороны по зрительному нерву в мозг с сетчатки глаза поступает лишь контурное их изображение, из-за оппонентной организации зрительных полей ганглиозных нейронов сетчатки [6]. Зрительный нерв содержит более одного миллиона волокон, которые являются аксонами (выходами) ганглиозных нейронов. По каждому волокну передаются сигналы с небольшой, приблизительно круглой области, локализации фоторецепторов сетчатки, причём ганглиозный нейрон суммирует сигналы рецепторов центра круга и вычитает из этой суммы сумму сигналов рецепторов периферии круга. Такая оппонентная структура описывается, например, двумерным оператором Марра ![]() [7], с учётом весовой функции суммирования. Вид весовой функции приведён на рисунке 7. Обнаружены [6] рецептивные поля с различным сочетанием трёх видов колбочек. Их обозначения представлены на рисунок 8. Здесь, кроме обозначений круглых рецептивных полей, приведены обозначения соответствующих масок (крестообразные фигуры) для оппонентно-цветового кодирования тестовых изображений при компьютерном моделировании естественного зрения.
[7], с учётом весовой функции суммирования. Вид весовой функции приведён на рисунке 7. Обнаружены [6] рецептивные поля с различным сочетанием трёх видов колбочек. Их обозначения представлены на рисунок 8. Здесь, кроме обозначений круглых рецептивных полей, приведены обозначения соответствующих масок (крестообразные фигуры) для оппонентно-цветового кодирования тестовых изображений при компьютерном моделировании естественного зрения.
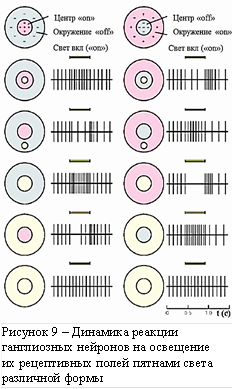
Рисунок 8 демонстрирует номенклатуру оппонентно-цветовых рецептивных полей, отличающихся не только сочетанием цветов, но и знаков (+) и (–), которые соответствуют сигналам возбуждения или торможения при активации рецепторов. Здесь красный цвет означает преимущественное действие в данной зоне рецептивного поля L-колбочек, зелёный – M-колбочек, синий – S-колбочек, жёлтый – как L, так и M-колбочек, белый – всех L, M, и S-колбочек.
Рецептивное поле любого нейрона зрительного пути можно определить, измеряя его сигнал при последовательном освещении рецепторов небольшими пятнами света различного цвета и диаметра круглой или кольцеобразной формы. Такое тонкое исследование с освещением колбочек сетчатки глаза, проводилось на обезьянах [6]. Реакции нейронов при различных воздействиях на их рецептивные поля показана на рисунке 9. Здесь слева обозначены реакции нейрона (осциллограммы потенциалов действия – спайков), который возбуждается при включении («on») света в центре его рецептивного поля, а справа реакции нейрона с другим типом рецептивного поля, возбуждающимся при выключении («off») света в центре. Реакции нейронов при аналогичном воздействии на периферию рецептивных полей соответствуют их торможению. Величина реакции пропорциональна частоте спайков справа от изображений рецептивных полей. Положение, размер и форма зоны освещения (жёлтой) изменяется по строкам изображения. Временной масштаб в правом нижнем углу позволяет сделать вывод о длительностях светового импульса и реакции нейрона. Можно видеть, что длительность реакции порядка одной секунды хорошо коррелирует со временем появления вышеописанного «пустого поля».
Обратное преобразование контурного изображения в изображение полноцветной раскраски возможно при наличии в зрительном тракте механизма восстановления изображения по его контурной версии и известным значениям яркости на его границах, что соответствует решению математической задачи Дирихле [8]. В пользу такого преобразования свидетельствует наличие особого оптического механизма равномерного освещения периферии сетчатки, открытого А. Л. Ярбусом [9].
Особым свойством зрительной системы человека является свойство цветовой константности, когда изменения спектра освещения не влечёт за собой существенного изменения цветности объектов наблюдения, что кажется парадоксальным, учитывая вышесказанное о форме спектра отражённого света. Далее рассмотрим возможный механизм обеспечения цветовой константности с учётом технологии восстановления контурного изображения.
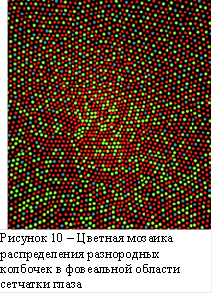
Кажется парадоксальным тот факт, что в центральной (фовеальной) области сетчатки глаза, соответствующей области ясного зрения, отсутствуют коротковолновые (S) колбочки, а присутствуют только средне- (M) и длинно- (L) волновые, и при этом мы ясно видим синие оттенки цвета. На рисунке 10 приведена мозаичная модель участка сетчатки глаза [10], которая показывает не только отсутствие синих элементов в центре рисунка, но и наличие довольно больших локальных скоплений здесь однородных по цвету элементов (особенно красных). Далее рассмотрим возможный механизм восприятия цвета с учётом движения глаз, устраняющий кажущийся парадокс.
Объясняя ранее отмеченные свойства зрительной системы, будем иметь в виду, что предложенные здесь их модели в технической области при создании искусственного интеллекта требуют разработки соответствующей элементной базы и необходимого программного обеспечения. В области физиологии зрения эти модели могут приниматься как гипотезы для обоснования проведения соответствующих исследований.